

地理隔离往往带来生殖隔离,作为人工物理屏障,长城是研究其对周围物种遗传分化影响的绝佳模型。研究人员从居庸关长城两侧的六个植物物种中各选择了一个种群,并从居庸关附近山顶的一条小路两侧的五个物种中各选择了一个种群。
材料和方法
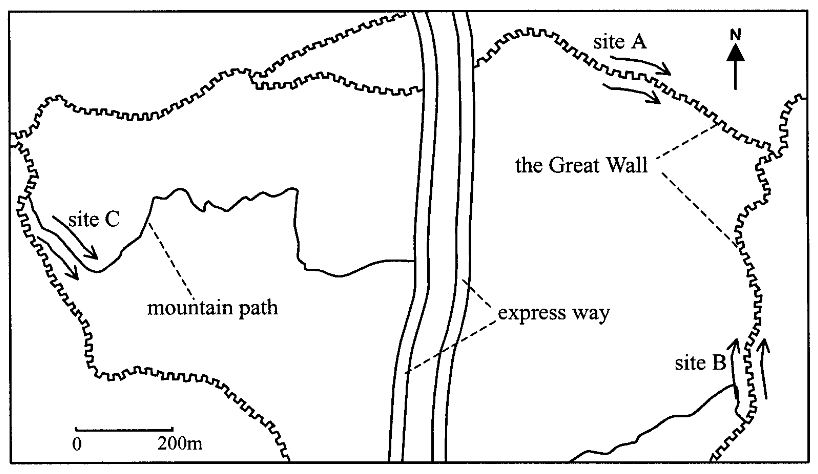
植物材料分为两次,1999年9月和2000年4月在居庸关长城的两个地点取样(A和B)和一条小路两侧的一个地点(地点C)采集
A和B实际上是两端长城的不同位置,C点的作用更类似于对照组
对收集到的嫩叶在室温环境下保存到硅胶中,使用CTAB方法从干燥叶片中提取总DNA。
RAPD bands were scored as present (1) or absent (0) for each DNA sample. The genetic diversity within sub- populations was measured by the percentage of poly- morphic bands, which was calculated by dividing the number of polymorphic bands by the total number of bands surveyed for the corresponding subpopulations (Table 2). Both Nei and Li’s and Euclidean genetic distances within populations were calculated employing RAPDistance version 1.04 (Armstrong et al, 1994). Based on Nei and Li’s genetic distance matrix, a dendrogram of individuals within a population was constructed using the neighbour-joining method. The variance components between subpopulations and among individuals within subpopulations were calculated with WINAMOVA ver- sion 1.55 (Excoffier, 1993) based on Euclidean genetic distances. The significance of each variance component was tested by a random permutation test. The signifi- cance of the difference in variance component between the Great Wall subpopulations and the control subpopu- lations was tested using a paired t-test.
每个 DNA 样本的 RAPD 条带分为存在(1)和不存在(0)。亚种群内的遗传多样性用多态性条带的百分比来衡量,多态性条带的计算方法是用多态性条带的数量除以相应亚种群的调查条带总数(表 2)。利用 RAPDistance 1.04 版(Armstrong 等,1994 年)计算种群内的 Nei 和 Li 遗传距离以及欧氏遗传距离。根据 Nei 和 Li 的遗传距离矩阵,使用邻接法构建了种群内个体的树枝图。根据欧氏遗传距离,用 WINAMOVA 1.55 版(Excoffier,1993 年)计算亚群之间和亚群内部个体之间的方差成分。每个方差分量的显著性通过随机排列测试进行检验。长城亚群与对照亚群之间方差分量差异的显著性采用配对 t 检验。
成果
居庸关长城沿线同一亚种群的个体往往聚集在一起,而沿线的一个种群的两个亚种群个体往往是混在一起。
居庸关长城地点亚群之间的分化程度明显高于对照地点,长城和沿岸的五个种群符合外交植物的遗传多样性范围。
超过71%的遗传变异归因于亚群内部的个体差异,虽然亚群之间的遗传变异差异较小,但被长城分隔的亚群之间的遗传差异较为明显。
被长城分隔的亚群之间的基因流受到了一定程度的干扰,尚不清楚亚群之间有限的基因流动是由风、动物还是人类活动造成。
同时还突出强调了风媒花榆树亚群的遗传分化程度要低于虫媒花榆树亚群,符合风媒花遗传分化程度要低于虫媒花。
在小种群中,遗传漂变可能在亚种群中发挥重要作用。
补充实验
为了排除不是长城,而是地形,诸如两边的光照、温度和湿度等环境因素不同对物种的形成的影响,研究人员在居庸关长城附近的山顶上选择了一个地理条件相似但是没有长城的地方。
尽管对照地点种群内部的亚群在遗传上有显著差异,但在衔接分析时,两个亚群中的所有个体都聚在一起,没有明显的亚群聚类,但是对照地点亚群之间的方差约为居庸关长城亚群之间的一般,长城地点的方差明显大于对照地点。
因此可以推断居庸关长城对植物亚群的基因流动起到了物理隔离的作用。
媒花遗传分化程度要低于虫媒花
- 传粉距离与基因流动: 风媒花依靠风力传播花粉,传粉距离往往较远。风可以携带花粉跨越很长的距离,这增加了不同个体间的基因交流(基因流动)。当种群之间的基因流动增加时,种群之间的遗传差异会降低,从而导致遗传分化程度较低。
相反,虫媒花依赖特定的昆虫或动物传粉,传粉距离相对较短,而且传粉通常局限于较小的范围。虫媒花的传粉往往具有一定的定向性,这限制了种群之间的基因流动,导致不同种群之间的遗传分化程度较高。
- 传粉特异性: 虫媒花常常与特定的昆虫或动物形成共生关系,这些传粉者可能具有一定的选择性,传粉范围相对较窄。虫媒花的这种特异性传粉会导致局部种群的遗传隔离,从而加速不同种群之间的遗传分化。
风媒花的花粉可以更随意地传播到大范围的其他个体,传粉者的选择性作用较小,基因交流更加广泛,因此种群之间的遗传差异不会像虫媒花那样显著。
- 花的形态与进化压力: 虫媒花为了吸引传粉者,往往进化出复杂的形态(如颜色、香气、花蜜等)来适应特定的传粉者。这样的共同进化关系可能会导致不同种群间的快速分化。而风媒花由于不依赖于特定的传粉者,一般花形较简单,不需要特殊适应性,这种进化压力相对较低,也减少了种群间的分化速度。
结论
被长城隔离的同种植物的亚居群间具有极为显著的遗传分化
被山顶小径隔开的亚居群间无显著的遗传分化
尽管只有几百年时间,长城作为一道人为的物理屏障,可能通过阻隔了植物的基因流动或造成了不同的微环境,从而对两侧植物居群的遗传分化发挥了作用